
Dutkowce
| Syringophilidae | |
| Lavoipierre, 1953 | |
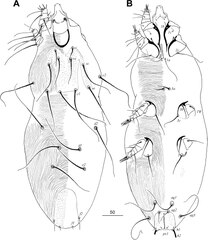 Tanopicobia trachyphoni (Picobiinae) | |
| Systematyka | |
| Domena | |
|---|---|
| Królestwo | |
| Typ | |
| Gromada | |
| Podgromada | |
| Nadrząd | |
| Rząd | |
| Podrząd | |
| Infrarząd | |
| Nadrodzina | |
| Rodzina |
dutkowce |
_-_Mites_-_Collection_Anthonie_Cornelis_Oudemans.jpeg)
_-_Mites_-_Collection_Anthonie_Cornelis_Oudemans.jpeg)
_-_Mites_-_Collection_Anthonie_Cornelis_Oudemans.jpeg)
Dutkowce[1] (Syringophilidae) – rodzina roztoczy z rzędu Trombidiformes i nadrodziny Cheyletoidea. Obejmuje ponad 320 opisanych gatunków, jednak ich całkowitą liczbę szacuje się na co najmniej 5 tysięcy. Są to obligatoryjne pasożyty ptaków, żyjące w dutkach ich piór, zwykle lotek. Odnotowano je na ponad 370 gatunkach ptaków z 22 rzędów. Mają robakowate, słabo zesklerotyzowane ciało o wydłużonej idiosomie, krótkich odnóżach i sztyletowatych lub harpunowatych szczękoczułkach. W rozwoju przechodzą przez stadia: trójnogiej larwy, protonimfy i tritonimfy. Rozród i rozwój odbywa się w dutce. Rodzina kosmopolityczna; zasięg geograficzny jej przedstawicieli odwzorowuje areał żywicieli.
Morfologia[edytuj | edytuj kod]
Postać dorosła[edytuj | edytuj kod]
Roztocze o wydłużonym, robakowatym ciele osiągającym od 500 do 1500 μm długości, ubarwionym mlecznobiało[2][3]. Dymorfizm płciowy jest silnie zaznaczony. Samce są bardziej krępej budowy i osiągają mniejsze rozmiary[3]. Ciało dutkowców podzielone jest na pseudotagmy – gnatosomę i idiosomę, a ta ostatnia z kolei na propodosomę i opistosomę, zwaną w tej grupie także hysterosomą[3][4].
Gnatosoma[edytuj | edytuj kod]
Gnatosoma jest przeciętnie wykształcona[4], wydłużona. Położone w jej brzusznej części[3] subkapitulum (infrakapitulum) jest osadzone głęboko w idiosomie i wyposażone w dobrze widoczną apodemę; na jego chetotaksję składają się szczecinki oznaczone jako elcp, n, ao1 i ao2[4]. Na szczycie gnatosomy leży stożkowaty hypostom o krawędzi wierzchołkowej zaokrąglonej, płaskiej, ściętej albo wyposażonej w rozmaitej wielkości wyrostki[3]. Hypostom zaopatrzony jest w wargi, a jego płaty przyrośnięte są do[4] położonego grzbietowo styloforu. U samic wielu gatunków hypostom ma parę przezroczystych guzków[4]. W środkowej, rostralnej części styloforu umieszczone są perytremy o kształcie litery „M” lub „U”[3][4]. Na gnatosomie osadzone są ponadto szczękoczułki i nogogłaszczki. Te pierwsze są długie, giętkie, równoległe, o formie sztyletów lub harpunów i w obrębie danego gatunku ich długość nie ulega zmianie w kolejnych stadiach rozwojowych[3]. Nogogłaszczki budują po cztery człony: krętarz, udo, kolano i stopogoleń[3][4]. Krętarz ma podstawę wchodzącą w brzuszną stronę idiosomy[4]. Na udzie występują szczecinki: d, v’, v”, na kolanie: d oraz l”, a na stopogoleniu: szczecinki: d, lT, l’, eupatidia: acm, sul, ul’, ul” oraz solenidion ω[4]. Wierzchołek stopogolenia w podrodzinie Picobinae jest ścięty, a u pozostałych dutkowców zaokrąglony[3].
Idiosoma[edytuj | edytuj kod]
Idiosoma jest całkowicie pozbawiona oczu i kupuli. Jej kompletna chetotaksja obejmuje szczeciny: scx, vi, ve, si, se, c1, c2, d1, d2, e2, f1, f2, h1, h2, ps1, ps2, ag1, ag2, ag3, g1, g2[4]. Propodosoma ma na stronie grzbietowej (propodonotum) w różnym stopniu zesklerotyzowaną, jednolitą lub podzieloną podłużnie tarczkę propodosomalną (propodonotalną)[3]. Szczeciny na wierzchu propodosomy występują w liczbie 5–6 par i zwykle mają formę włosowatą, rzadziej są pierzaste, spiralne lub pokryte guzkami, a ich rozmieszczenie jest charakterystyczne dla danego rodzaju[3].
Hysterosoma jest pseudotagmą dobrze rozwiniętą[4]. Na grzbietowej stronie hysterosomy (hysteronotum) typowo występuje tarczka hysterosomalna (hysteronotalna), która może być wtórnie rozdzielona na dwie tarczki, uwsteczniona, całkiem zanikła lub zrośnięta z leżącą na tyle hysterosomy tarczką pygidialną[3][5]. Tarczka pygidialna również bywa nieobecna[4]. Grzbietowa powierzchnia hysterosomy ma 5 par szczecin o kształcie włosowatym[3]. Otwory odbytowy i płciowy samicy umieszczone są na brzusznej stronie idiosomy lub na jej tylnym końcu i nakryte parą zmarszczek oskórka. U samca odbytu albo brak albo jest on połączony z otworem płciowym w jeden otwór płciowo-analny. Otwór ten umieszczony jest po grzbietowej stronie ciała. Edeagus jest rurkowaty, spiczasty[4], u szczytu może być rozdwojony. Szczeciny paragenitalne występują zwykle w liczbie trzech par położonych brzusznie[3]. Samce mają od 2 do 3 par szczecinek aggenitalnych, 2 pary szczecinek pseudanalnych i 2 pary szczecinek genitalnych, aczkolwiek te ostatnie mogą u niektórych gatunków wykazywać neotrichię[4].
W budowie przejawia się u Picobiinae polimorfizm samic w postaci nieczęstego pośród roztoczy zjawiska fizyogastrii[3][2]. W przeciwieństwie do samic typowych samice fizyogastryczne mają hystrosomę silniej wydłużoną, czasem paletkowato rozdętą, dzięki czemu mogą złożyć 2 lub 3 bardzo duże jaja, podczas gdy u pozostałych dutkowców samice składają od 10 do 12 mniejszych jaj[3].
Odnóża kroczne[edytuj | edytuj kod]
Idiosoma dysponuje czterema parami smukłych[4], dość krótkich[6] odnóży, z których dwie pierwsze leżą na propodosomie, a dwie tylne na hysterosomie[3][4]. Duża przerwa między nasadami odnóży drugiej i trzeciej pary należy do synapomorfii rodziny[7]. Odnóża składają się z sześciu członów: biodra, krętarza, uda, kolana, goleni i stopy zwieńczonej słabo wykształconym przedstopiem, które wyposażone jest w parę gładkich pazurków i pierzaste empodium[3][4]. Bywa, że dwie pierwsze pary odnóży są bardziej krępej budowy niż pozostałe. Na spodzie propodosomy epimeryty I pary mogą być równoległe względem siebie lub rozbieżne oraz mogą być wolne lub zrośnięte z epimerytami II pary[3]. Biodra pierwszej pary mają szczecinki: 1a, 1b, 1c, drugiej pary tylko szczecinkę 2c, trzeciej pary szczecinki: 3a, 3b, 3c, a czwartej pary tylko szczecinki 4b i 4c[4]; brak szczecinki 4a jest synapomorfią dutkowców[7]. Wszystkie krętarze mogą być wyposażone w szczecinkę brzuszną (v). Na udach dwóch pierwszych par odnóży mogą występować szczecinki grzbietowe i brzuszne (d i v), zaś na pozostałych ich parach tylko grzbietowe (d). Kolana pierwszej pary odnóży mają do trzech szczecinek o oznaczeniach: d, l’, σ, drugiej pary tylko szczecinki d i l’, a pozostałych par tylko szczecinkę l’. Na dwóch pierwszych parach goleni występują szczecinki: d, v, φ i para szczecinek l, na trzeciej parze szczecinka d i para szczecinek l, a na parze ostatniej tylko szczecinka d. Stopy pierwszej pary mają pojedyncze szczecinki ft i vs oraz parzyste szczecinki: tc, a, p, u. Druga para stóp ma pojedynczą szczecinkę vs oraz parzyste szczecinki: tc, p, u. Pozostałe pary stóp mają tylko trzy parzyste szczecinki: tc, p, u. Dwie początkowe pary stóp mają solnidia (ω), przy czym te na drugiej parze są umieszczone grzbietowo[4].
Postacie młodociane[edytuj | edytuj kod]
Postacie młodociane różnią się od dorosłych rozmiarami ciała, chetotaksją i sklerotyzacją oskórka, a ponadto postać larwalna pozbawiona jest odnóży czwartej pary. Większość gatunków ma stadia larwy i protonimfy pozbawione tarczek na propodonotum i hysteronotum. Tarczki te zwykle pojawiają się dopiero w stadium tritonimfy. Nogogłaszczki i propodonotum komplet szczecinek zyskują dopiero w stadium tritonimfy – u protonimf brakuje szczecinek h, vF oraz l”G, a u larwy także szczecinki l”F. U gatunków wykazujących neotrichię larwy pozbawione są szczecinek aggenitalnych, protonimfy mają ich tylko jedną parę, tritonimfy dwie pary, a trzecia para pojawia się u formy dorosłej. U gatunków wykazujących hipertrichię szczecin aggenitalnych cecha ta pojawia się w stadium tritonimfy, podczas gdy larwa i protonimfa wyglądają jak u gatunków z neotrichią. U większości gatunków larwy i protonimfy mają po jednej parze szczecinek genitalnych i pseudanalnych, tritonimfy mają dwie pary szczecinek pseudanalnych i jedną genitalnych, a formy dorosłe po dwie pary szczecinek genitalnych i pseudanalnych. Pola koksalne (pozostałości bioder) larw są pozbawione szczecinek z wyjątkiem pierwszej ich pary, gdzie obecne są szczecinki 1b'. U protonimf następujące szczecinki pojawiają się na trzech początkowych parach pól: 1c, 2c, 3b i 3c. Czwarta para pól zyskuje szczecinki 4b i 4c w stadium tritonimfy. Krętarze są pozbawione szczecinek aż do stadium tritonimfy, w którym to szczecinki l’R pojawiają się na ich trzech pierwszych parach. Kolana larw i protonimf mają po jednej grzbietowej i jednej bocznej szczecince na dwóch początkowych parach oraz jednej bocznej szczecince na parze trzeciej. Szczecinka boczna czwartej pary kolan pojawia się w stadium tritonimfy. Larwy mają trzecią parę stóp pozbawioną szczecinek wachlarzowatych. U protonimf takowe pojawiają się na trzeciej parze stóp, ale brak ich na parze czwartej, która zyskuje je dopiero w stadium tritonimfy[2].
Biologia i ekologia[edytuj | edytuj kod]
Wszyscy przedstawiciele rodziny są obligatoryjnymi pasożytami zewnętrznymi ptaków. Bytują wewnątrz dutek ich piór. Żerują na płynie tkankowym, przebijając w tym celu ścianę dutki swymi sztyletowatymi lub harpunowatymi palcami ruchomymi szczękoczułków[6][3]. Nowo wykształcające się dutki zajmowane są przez roztocze za pośrednictwem ich naturalnego otworu – umbilicus superiorus[6]. Większość przedstawicieli rodziny wyspecjalizowana jest w zamieszkiwaniu lotek. Tylko przedstawiciele Picobiinae występują w dutkach opierzenia ciała[4].
Obie płcie przechodzą w rozwoju przez stadia larwy, protonimfy i tritonimfy; tak jak u innych przedstawicieli nadrodziny brak w ich rozwoju stadium deutonimfy[4][2]. Wylinka następuje przez poprzeczne pęknięcie oskórka w pobliżu bruzdy dysjugalnej (linienie typu merodehiscenicznego)[2]. Rozwój i rozród odbywa się wewnątrz dutki. Samce spędzają w niej całe życie. Tylko dorosłe, zapłodnione samice opuszczają swoją dutkę celem zajęcia nowych[6].
Do dutkowców należą głównie pasożyty mono- i oligokseniczne. Większość gatunków ograniczona jest do jednego gatunku lub rodzaju żywiciela, a przypadki występowania jednego gatunku u ptaków z różnych rodzin czy rzędów należą do rzadkości[6]. Do 2015 roku dutkowce stwierdzono na 482 gatunkach ptaków z 95 rodzin i 24 rzędów[8]. Na tym samym gospodarzu występować mogą przedstawiciele różnych rodzajów, a nawet podrodzin dutkowców[9][6][4], jednak przypadki zasiedlenia tej samej dutki przez więcej niż jeden gatunek należą do rzadkości[4].
Rozprzestrzenienie[edytuj | edytuj kod]
Takson kosmopolityczny, znany ze wszystkich krain zoogeograficznych oprócz Antarktydy[8]. Zasięg poszczególnych gatunków pokrywa się najpewniej z zasięgiem ich żywicieli, jednak ich rozmieszczenie jest niedostatecznie zbadane[3]. W Polsce do 2015 roku stwierdzono występowanie 66 gatunków[8].
Taksonomia[edytuj | edytuj kod]
Takson ten wprowadzony został w 1953 roku przez Michela M.J. Lavoipierre’a dla objęcia monotypowego wówczas rodzaju Syringophilus, zaliczanego wcześniej do Myobiidae[10]. Niezależnie utworzenia rodziny Syringophilidae dla rodzajów Syringophilus i Picobia dokonał w 1957 roku Wsiewołod Dubinin[11][6].
Dutkowce umieszcza się w nadrodzinie Cheyletoidea. Zarówno dane morfologiczne jak i analizy molekularne wskazują, że dutkowce zajmują w jej obrębie pozycję siostrzaną względem Cheyletidae[7][12][6].
Pierwszej dużej rewizji rodziny dokonał w 1970 roku John Kethley[9]. Trzy lata później opublikował on wspólnie z Donaldem E. Johnstonem wyniki analizy fenetycznej, na podstawie której autorzy podzielili rodzinę na dwie podrodziny: Syringophilinae i Picobiinae[13]. W 1977 roku Stanley D. Casto wyróżnił w ich obrębie kolejną podrodzinę – Loabatinae[14], jednak jej jedyny rodzaj włączony został do Picobiinae w 2000 roku przez Alexandra Faina, Andre Bochkova i Siergieja Mironowa[15]. Pierwszej, morfologicznej, analizy filogenetycznej rodziny dokonali w 2013 roku Maciej Skoracki, Eliza Głowska i Andre Bochkov. O ile Picobiinae w jej wynikach były monofiletyczne, tak Syringophilinae okazały się względem nich parafiletyczne[6].
Do 2014 roku opisano ponad 330 gatunków dutkowców[16] (334 według checklisty z 2015[8]), zgrupowanych w 62 rodzajach[17][16][8]. Ogólną liczbę ich gatunków ocenia się jednak na nie mniej niż 5 tys.[3][2] Należą doń m.in.[6][8][4]:
- Apodisyringiana Skoracki, 2005
- Apodisyringiophilus Skoracki et OConnor
- Ascetomylla Kethley, 1970
- Aulobia Kethley, 1970
- Aulonastus Kethley, 1970
- Betasyringophiloidus Skoracki, 2011
- Blaszakia Skoracki et Sikora, 2008
- Bochkovia Skoracki et OConnor, 2010
- Bubophilus Philips et Norton, 1978
- Calamincola Casto, 1978
- Castosyringophilus Bochkov et Perez, 2003
- Charadriineopicobia Skoracki, Spicer et OConnor
- Charadriphilus Bochkov et Chistyakov, 2001
- Chenophila Kethley, 1970
- Ciconichenophilus Skoracki et OConnor, 2010
- Colinophilus Kethley, 1973
- Colisyringophilus Skoracki, Unsoeld et Ozminsk
- Columbiphilus Kivganov et Sharafat
- Corvitorotroglus Skoracki et Bochkov
- Creagonycha Kethley, 1970
- Crotophagisyringophilus Skoracki, 2008
- Cuculisyringophilus Skoracki, 2005
- Fritschisyringophilus Bochkov, Fain et Skoracki, 2004
- Galliphilopsis Skoracki et Sikora 2004
- Gunabopicobia Skoracki et Hromada
- Ixobrychiphilus Skoracki, Zmudzinski et Solarczyk, 2017[16]
- Kalamotrypetes Casto, 1980
- Kethleyana Kivganov, in Kivganov et Sharafat 1995
- Krantziaulonastus Skoracki, 2011
- Lawrencipicobia Skoracki et Hromada
- Megasyringophilus Fain, Bochkov & Mironov, 2000
- Meitingsunes Glowska et Skoracki, 2010
- Mironovia Chirov & Kravtsova, 1995
- Neoaulobia Fain, Bochkov & Mironov, 2000
- Neoaulonastus Skoracki, 2004
- Neopicobia Skoracki, 2011
- Neoperisterophila Skoracki, 2005
- Neosyringophilopsis Skoracki et Sikora
- Niglarobia Kethley, 1970
- Paraniglarobia Skoracki
- Peristerophila Kethley, 1970
- Phalarophilus Skoracki, Bochkov et OConnor, 2011
- Phipicobia Glowska et Schmidt
- Philoxanthornia Kethley, 1970
- Picineoaulonastus Skoracki, Klimovičová, Muchai et Hromada
- Picisyringophilus Skoracki et OConnor, 2010
- Picobia Haller, 1878
- Procellariisyringophilus Schmidt et Skoracki, 2007
- Pseudopicobia Skoracki, Scibek et Sikora
- Psittaciphilus Fain, Bochkov et Mironov, 2000
- Pteroclidisyringophilus Skoracki, 2011
- Rafapicobia Skoracki, 2011
- Selenonycha Kethley, 1970
- Stibarokris Kethley, 1970
- Syringophiloidus Kethley, 1970
- Syringophilopsis Kethley, 1970
- Syringophilus Heller, 1880
- Terratosyringophilus Bochkov et Perez
- Tinamiphilopsis Skoracki et Sikora, 2004
- Torotrogla Kethley, 1970
- Trypetoptila Kethley, 1970
- Terratosyringophilus Bochkov et Perez, 2003
Przypisy[edytuj | edytuj kod]
- ↑ Maciej Skoracki: Dutkowce. Syringophilidae. W: Fauna Polski. Charakterystyka i wykaz gatunków. Tom III. Wiesław Bogdanowicz, Elżbieta Chudzicka, Irmina Pilipiuk, Ewa Skibińska (red.). Warszawa: Muzeum i Instytut Zoologii PAN, 2008, s. 131.
- ↑ a b c d e f Maciej Skoracki, Quill mites (Acari: Syringophilidae) of the Palaearctic region, „Zootaxa”, 2840 (1), 2011, s. 1–415, DOI: 10.11646/zootaxa.2840.1.1, ISSN 1175-5334.
- ↑ a b c d e f g h i j k l m n o p q r s t u v Budowa morfologiczna ektopasożytniczych roztoczy zrodziny Syringophilidae (Acari: Prostigmata): Stawonogi. Interakcje pasożyt–żywiciel. Alicja Buczek, Czesław Błaszak (red.). Lublin: Liber, 2004, s. 19-22.
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x M. Skoracki, S.A. Zabludovskaya, A.V. Bochkov. A review of Prostigmata (Acariformes: Trobidiformes) permanently associated with birds. „Acarina”. 20 (2), s. 67–107, 2012.
- ↑ Maciej Skoracki. Quill mites of the genus Syringophilopsis (Acari, Syrin-gophilidae) from passeriform birds of Poland with descriptions of five new species. „Acta Parasitologica”. 49, s. 45-62, 2004.
- ↑ a b c d e f g h i j Maciej Skoracki, Eliza Głowska, Andre V. Bochkov. Phylogeny of quill mites of the family Syringophilidae (Acari: Prostigmata) based on their external morphology. „European Journal of Entomology”. 110 (4), s. 663–675, 2013. DOI: 10.14411/eje.2013.090. ISSN 1210-5759.
- ↑ a b c A.V. Bochkov. New observations on phylogeny of cheyletoid mites (Acari: Prostigmata: Cheyletoidea). „Proceedings of the Zoological Institute RAS, St. Petersburg”. 312, s. 54–73, 2008.
- ↑ a b c d e f E. Głowska, M. Chrzanowski , K. Kaszewska. Checklist of quill mites (Acariformes: Syringophilidae) of the World. „Zootaxa”. 3968, s. 1-81, 2015.
- ↑ a b J.B. Kethley. A revision of the family Syringophilidae (Prostigmata: Acarina). „Contrib. Am. Entomol. Inst.”. 6, s. 1–76, 1970.
- ↑ M.M.J. Lavopierre. The undescribed male and female of the Pigeon quill mite, Syringophilus columbae Hirst, 1920. „Trans. R. Soc. Trop. Med. Hyg.”. 47 (7), 1953.
- ↑ W.B. Dubinin. A new classification of the mites of the superfamilies Cheyletoidea and Demodicoidea (Acariformes, Trombidiformes). „Parasitol. Sb. Acad. Nauk SSSR”. 17. s. 17–136.
- ↑ Mirosława Dabert, Wojciech Witaliński, Andrzej Kaźmierski, Ziemowit Olszanowski. Molecular phylogeny of acariform mites (Acari, Arachnida): Strong conflict between phylogenetic signal and long-branch attraction artifacts. „Molecular Phylogenetics and Evolution”. 56 (1), s. 222-41, 2010. DOI: 10.1016/j.ympev.2009.12.020.
- ↑ Donald E. Johnston, John B. Kethley. A numerical phenetic study of the quill mites of the family Syringophilidae (Acari). „J. Parasitol.”. 59, s. 520–530, 1973.
- ↑ S.D. Casto. Cuculiphilus lobatus gen. n., sp. n. representing a new subfamily of quill mites (Acarina: Syringophilidae) from the groove-billed ani, Crotophaga sulcirostris (Cuculiformes: Cuculidae). „Southw. Nat.”. 22, s. 169–176, 1977.
- ↑ A. Fain, A.V. Bochkov, S.V. Mironov. New genera and species of quill mites of the family Syringophilidae (Acari: Prostigmata). „Bull. Inst. R. Sci. Nat. Belg.”. 70, s. 33–70, 2000.
- ↑ a b c Skoracki, Maciej; Zmudzinski, Mateusz; Solarczyk, Piotr. Ixobrychiphilus, a new genus of the family Syringophilidae (Acariformes: Prostigmata). „Acarologia”. 57 (2), s. 269-273, 2017.
- ↑ Miroslava Klimovičová, Martin Hromada. New hosts and localities of quill mites (Acari: Syringophilidae) parasitising birds in Slovakia. „Acta Universitatis Prešoviensis. Folia Oecologica”. 11, s. 31-42, 2014.